MN和Ss血型的N末端基本相同,你了解吗?
时间:作者:起名专家
MN和Ss血型的N末端基本相同,你了解吗?
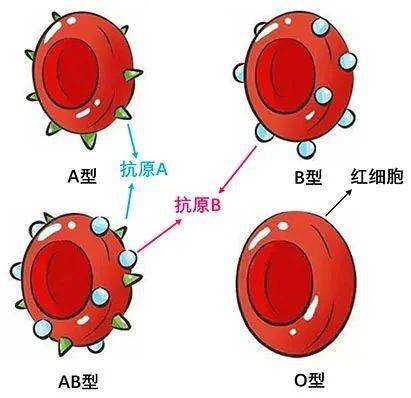
MN 抗原位于 GYPA 上,GYPA 是最丰富的红细胞糖蛋白之一。M 和 N 抗原是由 GYPA 的前 5 个氨基酸编码的 2 个常染色体共显性抗原,包括 3 个 O-连接聚糖作为表位的一部分。M 和 N 在氨基酸 1 和 5 处不同,其中 M 是 ser-ser-thr-thr-gly,N 是 leu-ser-thr-thr-glu。M 是祖先的 GYPA 等位基因,在所有人类种群和旧大陆猿中都很常见。GYPA、血型糖蛋白 B(GYPB;)和血型糖蛋白 E(GYPE;)在染色体 4q31 上紧密相连。GYPB 的 N 末端与 GYPA 的 N 末端基本相同,只是它始终表达 N 抗原,表示为“N”或 N-prime。Ss 血型的抗原( ) 位于 GYPB 上MN和Ss血型的N末端基本相同,你了解吗?,GYPA、GYPB 和 GYPE 之间的重组和基因转换导致杂合血型糖蛋白分子和低发生率抗原的产生。因此,MN 和 Ss 血型统称为 MNSs 或 MNS 血型系统。U 抗原是指 GYPB 中位于膜附近的一个短的细胞外序列。重组导致 3 种血型糖蛋白无效表型: 由于 GYPA 和 GYPB 之间的重组,En(a-) 细胞缺乏 GYPA;由于 GYPB 中的重组,GYPB 阴性(SsU-) 细胞缺乏 GYPB;由于 GYPA 和 GYPE 之间的重组,M(k) 细胞(MNSsU-) 缺乏 GYPA 和 GYPB。具有血型糖蛋白无效表型的个体唾液酸含量降低,对疟疾感染的抵抗力增加(见)。GYPA 和 GYPB 对红细胞发育或存活不是必需的,并且 GYPA 和 GYPB 无效表型与贫血或红细胞功能改变无关(评论,2015 年)。
▼ 测绘
------
通过使用 cDNA 探针的原位杂交, 等人(1987)将 GYPA 基因定位到染色体 4q28-q31。
通过原位杂交和 RFLP 研究,在染色体 2 和 4 之间平衡的从头易位的情况下, 等人(1989)得出结论,纤维蛋白原基因簇( ) 位于 GYPA/GYPB 基因座的近端,并且所有这些基因座都位于 4q28 带中。
Wakui 等人在患有 4q 从头间质缺失的畸形女婴中(1991)发现 MN 基因座是完整的。根据这一发现和先前的映射数据(见历史),他们得出结论,MN 基因座位于 4q28.2-q31.1 段。
Onda 和 (1995)报道 GYPA、GYPB 和 GYPE 基因簇跨越约 330 kb 的染色体 4q31。
▼ 分子遗传学
------
和 (1978)发现 MM 血型多肽在 2 个氨基酸上与 NN 多肽不同MN和Ss血型的N末端基本相同,你了解吗?,它们是 MM 中的丝氨酸和甘氨酸以及 NN 中的亮氨酸和谷氨酸。MN 个体显示所有 4 个氨基酸。人红细胞膜的 2 种主要唾液酸糖蛋白 α 和 δ(分别为血型糖蛋白 A 和 B)具有 MNS 抗原特异性。它们的 N 末端的前 26 个残基具有相同的氨基酸序列。α 表示 M 或 N 血型活性;δ 只携带 N 型血的活性。此外,α 位置 26 处的天冬酰胺携带一条寡糖链,该链在 δ 的相同位置不存在。2 种唾液酸糖蛋白的剩余氨基酸序列不同,δ 表达 Ss 活性。
Mawby 等人使用针对主要唾液酸糖蛋白 α 不同结构区域的抗体(1981)证实 2 种变体形式( V 类和 Ph)代表了由编码 α 和 δ 的基因之间的异常交叉事件引起的杂合唾液糖蛋白分子。这些基因似乎是紧密相连的,顺序是 α-δ(5-prime 到 3-prime)。
具有罕见 En(a-) 变体的红细胞对恶性疟疾具有抗性( 等人,1982 年)。这种细胞缺乏血型糖蛋白 A,主要的红细胞唾液糖蛋白( 和mn血型遗传方式,1986)。Ss 系统的罕见 U(-) 变体缺乏其他主要唾液酸糖蛋白血型糖蛋白 B,对入侵具有相对抵抗力。Wr(b) 阴性细胞也能抵抗恶性疟原虫的入侵,尽管它们的表面具有正常量的糖蛋白 A 和 B。所有这些观察结果,以及使用抗血型糖蛋白和某些糖类(特别是 N-乙酰氨基葡萄糖)抗体的实验,都导致了血型糖蛋白在恶性疟原虫红细胞侵袭中的作用的初步模型( 和 ,1982)。
朗洛伊斯等人(1986)研究了血型糖蛋白 A 基因座(GPA) 表达缺失的红细胞频率。每个红细胞大约有 500,000 个拷贝的糖蛋白 A。GPA 的 2 种等位基因形式,血型 M 和血型 N,除了在氨基末端的 1 和 5 位有 2 个氨基酸取代外,是相同的( 等,1981)。 等人使用单克隆抗体(1986)鉴定了表达缺失突变体。他们发现,在暴露于诱变化疗药物后,正常人的变异细胞频率约为 100,000 个细胞中的 1 个,而癌症患者的变异细胞显着增加。
朗洛伊斯等人(1987)证明了血型糖蛋白 A 基因座的突变频率与原子弹幸存者的辐射暴露之间存在线性关系。
拉韦尔等人(1988)通过对来自正常 En(a+) 和罕见 En(a-) 人的基因组 DNA 的 印迹分析表明,血型糖蛋白 A 基因具有复杂的组织结构,并且在 En(a-) 表型的人中大量缺失(芬兰型),他们的红细胞中缺乏血型糖蛋白 A。拉韦尔等人(1988)得出结论mn血型遗传方式,芬兰的变异是纯合的,因为血型糖蛋白 A 基因完全缺失,而编码血型糖蛋白 B 或 C 的基因没有任何可检测的异常(1988) 确定了血型糖蛋白 A 和 B 基因的几个异常,导致他们得出结论,两者都被大量删除,被由血型 M 型血型糖蛋白 A 的 N 端部分和 C 端部分组成的基因融合产物取代血型糖蛋白 B 的一部分。
大久保等人(1988)描述了 2 个日本姐妹的近亲父母显然是 M(k) 纯合子。在其中 1 个姐妹中证明了红细胞膜中完全不存在唾液酸糖蛋白 A(α) 和 B(δ)。这是第三个报道的家庭;其他家庭之一也是日本人。除了本研究中患有霍奇金病的提议者外,所有受影响的个体都是健康的。
黄等人(1988)研究了一个家族,其中 3 个不同的血型糖蛋白突变存在于 16 名成员家族的 2 个个体中。变体 Dantu 血型糖蛋白显示出与 δ-α(GPB/GPA) 杂合血型糖蛋白一致的特性。该基因与编码 M 特异性 α 血型糖蛋白的基因有关。另一种变异型血型糖蛋白 Mi-III 型血型糖蛋白作为常染色体显性性状遗传,并与 N 血型活动相关。遗传模式表明它可能是 δ 血型糖蛋白(血型糖蛋白 B)的变体。在同时具有 Dantu 和 Mi-III 糖蛋白的人中,观察到 δ 糖蛋白缺乏,这表明 δ 基因的缺失或改变可能与 Dantu 基因存在于同一条染色体上。黄等人(1989) 表明 St(a)(Stone) 抗原同样由血型糖蛋白 A 和 B 基因的融合杂交体决定。
如前所述,血型糖蛋白变体 V 类分子(MiV) 是一种杂交体:Kudo 等(1990)表明基因的 5-prime 一半来自 GPA 基因,而 3-prime 一半来自 GPB 基因。这种结构与血型糖蛋白变体 St(a) 互惠,后者具有 GPB-GPA 混合结构。黄等人(1992)确定了在观察到第一个纯合子的白色家族中导致 I 类(MiI) 表型变化的分子性质。
St(a) 抗原已被证明与几种血型糖蛋白同工型有关。St(a) 等位基因与 GPA 基因中的剪接突变或 GPA 和 GPB 基因之间的杂交形成在遗传上相关。黄等人(2000)报道了第一次也是罕见的基因转换事件,其中 GPE 与 GPA 重组,产生了编码 St(a) 抗原的新型 GPA-EA 杂合基因。
和 Huang(1995)回顾了血型糖蛋白基因家族 25 种变体的分子遗传学,其共同点是它们产生于不等的基因组合或基因转换与剪接位点突变相关联。大多数重排发生在 2-kb 区域内,主要发生在 GPA 和 GPB 内,很少发生在第三个成员 GPE 内。他们观察到,关键特征是 2 个特定外显子(其中 1 个是沉默的)内的序列改组,这些外显子在 2 个亲本基因中是同源的。这导致该区域内序列嵌合体的表达,导致多态性。
MNSs 血型系统和疟疾
具有罕见 En(a-) 变体的红细胞对恶性疟(见)具有抗性( 等,1982)。这种细胞缺乏血型糖蛋白 A,主要的红细胞唾液糖蛋白( 和 ,1986)。Ss 系统的罕见 U(-) 变体缺乏其他主要唾液酸糖蛋白血型糖蛋白 B,对入侵具有相对抵抗力。Wr(b) 阴性细胞也能抵抗恶性疟原虫的入侵,尽管它们的表面具有正常量的糖蛋白 A 和 B。所有这些观察结果,以及使用抗血型糖蛋白和某些糖类(特别是 N-乙酰氨基葡萄糖)抗体的实验,都导致了血型糖蛋白在恶性疟原虫红细胞侵袭中的作用的初步模型。帕斯沃尔和威尔逊,1982 年)。
GYPA 和 GYPB 分别决定了 MN 和 Ss 血型,它们是在红细胞表面表达并与恶性疟原虫配体相互作用的 2 种主要受体。柯等人(2011)分析了 15 个不同疟疾暴露水平的非洲人群中血型糖蛋白基因家族的核苷酸多样性。在这些基因中发现了高水平的核苷酸多样性和基因转换。柯等人(2011)观察到这些重复基因之间以及 GYPA 不同细胞外结构域之间的遗传变异的不同模式。具体来说,他们确定了 GYPA 外显子 3 到 4 的固定适应性变化。相比之下,Ko 等人(2011)在许多人群中观察到 GYPA 外显子 2 的等位基因频谱偏向显着过量的中频等位基因;频谱失真的程度与疟疾暴露有关,可能是因为基因转换和平衡选择的共同影响。柯等人(2011)还发现了一种导致血型糖蛋白 B 胞外域发生 3 个氨基酸变化的单倍型。这种单倍型可能在 5 个高度接触疟疾的人群中适应性进化。
通过分析来自人群的基因组序列数据, 等人(2017)确定了一系列影响 GYPA 和 GYPB 的大拷贝数变异。他们发现,涉及 GYPB 丢失和 2 个 GYPB-GYPA 杂合基因编码 MNS 血型系统丹图抗原的复杂结构重排解释了附近区域与严重疟疾保护之间的关联。与3个基因(GYPE、GYPB和GYPA)的参考单倍型相比,保护性单倍型具有5个GYP基因,包括2个GYPE拷贝、2个Dantu杂种基因和1个GYPA拷贝。保护性单倍型在肯尼亚地区将严重疟疾的风险降低了 40%,但尚未在西非发现。
卡里乌基等人(2020)证明了丹图对疟原虫恶性疟原虫的裂殖子形式侵入红细胞的能力的影响。卡里乌基等人(2020)发现 Dantu 与红细胞表面发现的蛋白质库的广泛变化有关,但出人意料的是,入侵的抑制与特定的红细胞(RBC)-寄生虫受体-配体相互作用无关。通过使用视频显微镜跟踪入侵, 等人(2020)发现 RBC 张力与裂殖子入侵之间存在密切联系,并确定了一个张力阈值,高于该阈值很少发生入侵,即使在非 Dantu RBC 中也是如此。Dantu RBC 比非 Dantu RBC 具有更高的平均张力,这意味着更大的比例抵抗入侵。卡里乌基等人(2020)得出的结论是,他们的研究结果既解释了丹图的保护作用,又提供了新的见解,即为什么在个体内部和个体之间发现的异质红细胞群中,恶性疟原虫入侵的效率可能会有所不同。
GYPA 变异分析
Grant 和 (1994)讨论了使用 GPA 分析来评估癌症化疗产生的体细胞突变。
罗斯曼等人(1995)使用 GPA 测定来评估职业接触苯的影响。GPA 测定测量在杂合(MN) 个体的血液样本中失去 M 型血表达的变异红细胞的频率。通过用对 M 和 N 形式特异的荧光标记的单克隆抗体处理球形固定红细胞,并通过流式细胞术,对结合抗 N 抗体但不结合抗 M 抗体的变体细胞进行计数来检测变异细胞。变异细胞具有表型 N-0(单拷贝表达 N,不表达 M)或 NN(双拷贝表达 N,不表达 M)。这些表型变异源于前体细胞的不同突变机制:N-0 细胞被认为源于点突变、缺失或基因失活,罗斯曼等人(1995)使用这种 GPA 测定来评估中国上海 24 名重度暴露工人和 23 名匹配对照中苯造成的 DNA 损伤。在接触苯的工人中发现 MN GPA 变异细胞频率显着增加,但对于 N-零细胞,两组之间不存在显着差异。此外,终生累积职业接触苯与 NN 频率相关,但与 N 零频率无关,这表明 NN 突变发生在寿命较长的骨髓干细胞中。
▼ 历史
------
根据对易位染色体儿童家庭的研究, 等人(1968)建议 MN 基因座要么位于 2 号染色体的中间,要么靠近 4 号染色体长臂的远端。使用“条带技术”, 和 (1973)重新研究了他们在 1968 年报告的易位并得出结论MN 可以暂时分配到 2 号染色体长臂近端部分 q14 带的区域。 等人(1972)提供的数据表明 MN 基因座和 β 血红蛋白基因座( ) 是相关联的(当然,这已被反驳。) 等人(1975)排除了 MN 和 Hb β 小于 0.30 的重组分数。结果支持雄性较低的重组分数。怀疑与阿尔茨海默病基因座( ) 和结肠息肉病( ) 相关。重组数据表明 MN 和酸性磷酸酶(ACP1; ) 基因座相距很远( 等,1975)。库克等人(1978)通过排除映射将 MNS 从 9 号染色体排除,该排除映射结合了来自具有染色体标记的家族和具有牢固分配的标记的连锁研究的数据。MNSs 随后被分配到 4 号染色体。在对 2q;4q 易位家族的进一步研究中, 等人(1979)通过条带显示中断发生在 2q14 和 4q29,并且在中断部位丢失了一个分钟片段。
丢失是来自 2 号染色体还是 4 号染色体尚不确定,因为两者在这些位点都有几个短带,而先证者中只有一个带缺失。先证者缺乏他应该从他的 ss 父亲那里得到的血型,有改良的红细胞膜的迹象,并且有发育异常。由于表型异常与染色体异常同时出现, 等(1979)建议删除是所有更改的基础。由于(1978)报告的观察结果强烈表明 MNS 不接近 2q14, 等人(1979)得出结论,它必须在 4q29 附近的带中。库克等人(1980)比 4q31 更喜欢 4q28。对于雄性,Bias 和 (1979)发现 ( ) 和 MNS 的连锁在 theta 0.18 处的最大对数值为 3.99 。酸性磷酸酶和 Kidd 对 的 lods 均为 0.32,男性-θ 为 0.20。(1978)排除了 Gc 和 MNS 在男性中低于 25% 和女性中低于 30% 的重组频率的连锁。对于 MN 与 Gc,Falk 等人(1979)发现重组分数为 0.30 时,男性 lod 得分为 3.75。在雌性中,重组分数为 0.42 时的最大 lod 得分为 0.34。通过对 4 号染色体重排家族的 MNS 血型分析,包括缺失分析和家族连锁研究,Cook 等人(1981)得出结论,MNS 位点位于 4q28-q31 区域。Gedde-Dahl 和 (1981)建议序列可能是 MN--Ss--Gc。
在前 RFLP 时代,人类中测量的最长遗传间隔之一是 GC 和 MN 之间的间隔mn血型遗传方式,男性的 lod 得分为 3.79,重组分数为 0.32(Falk,1984)。在 MN 和 Ss 的 146 个信息家族的连锁分析中, 等人(1984)在 467 名重组儿童中发现了 7 名重组儿童,其中 1 名已确认重组(重新测试且与 HLA 兼容)和 6 名未验证。重组估计值的 95% 置信区间为 0.0033-0.1167。
由起名专家专业的命理起名平台。